
© elfruler 2023
PHOTOPERIOD, CIRCADIAN RHYTHMS, and CIRCANNUAL RHYTHMS
Most viewers of Bald Eagle nest cams are aware of how important photoperiod, circadian rhythms, and circannual rhythms are in the lives of birds. They are fundamental to how eagles’ behaviors and physiological processes engage with the environment around them. This article provides information about what these systems are and how they work, and it explores some of the complexities of how they govern the processes of the Life History of Bald Eagles.
The photoperiod is the timespan of sunlight within a 24-hour day, usually called daytime or daylight. Its counterpart is the scotoperiod, the timespan of darkness, or nighttime. As everyone knows, the photoperiod and scotoperiod change from one day to the next following an annual cycle, and they also vary by latitude (location on the earth north or south of the equator). Changes in photoperiod and the seasons of the year are caused by the Earth’s tilt on its axis by about 23.5°and its revolution around the sun every 365.4 days. In the Northern Hemisphere during the winter months the North Pole tilts away from the sun, making the sun lower in the sky and the daytimes short. During summer the North Pole tilts toward the sun, which thus is higher in the sky and the daytimes are long.
-
- The photoperiod is shortest on the Winter Solstice (~ December 21) (Lat. sol = sun + status = standing still) when the sun is lowest in the sky.
- The photoperiod slowly increases through the Vernal (Spring) Equinox (~ March 20) (Lat. equus = equal + nox = night) when the Earth’s axis is tilted neither toward nor away from the sun, and day and night are of roughly equal duration.
- The photoperiod continues to increase until it reaches its maximum at the Summer Solstice (~ June 20) when the sun is highest in the sky.
- It then slowly decreases through the Autumnal Equinox (~ September 22) when the axis again is not tilted in relation to the sun, and day and night are of roughly equal duration.
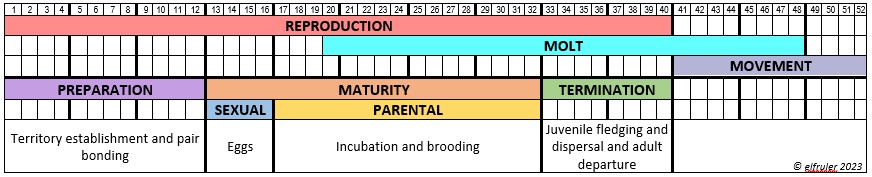
This progression is mirrored in the annual cycle—or as ornithologists call it the Life History—of most birds, including Bald Eagles. Each species has its own Life History, which is part of its genetic make-up, a long-term adaptation to ensure its survival. (See more discussion of Life History here.) The central Life-History stage for birds is Reproduction, which obviously is necessary for all species’ survival. This is followed by Molt, the regular replacement of feathers, which also is essential. Many species then go on Migration, or, as I prefer to call it in reference to Bald Eagles, a post-breeding stage of Movement. Here is a schematic diagram of the Life-History stages of Bald Eagles, mapped onto 52 weeks of the year, showing the typical durations of each stage. Notice that Molt overlaps with Reproduction at the beginning and with Movement at the end (click on the image for an enlarged view):![]()
The Life-History stages for a species revolve around the time of the year when optimal food resources are readily available to feed growing chicks as well as the parents who nurture them. Scientists refer to food availability as the ultimate factor that is critical to the timing of a species’ Reproduction. This, of course, is closely linked with the changes of seasons caused by the yearly movements of the Earth. Birds are able to “read” the seasons so that they move through the Life-History stages in such a way as to optimize their breeding success. They do this by means of internal rhythms (endogenous), biological clocks throughout an organism’s cells (which all organisms have) and that are correlated with the external rhythms (exogenous) of the changing photoperiod.
-
- Circannual rhythms ( circa = about and annum = year) are months-long cycles that govern a bird’s Life History.
- Circadian rhythms (Lat. circa = about + dies = day) are daily oscillations between high and low physiological activity.
Circannual rhythms and circadian rhythms are built into a bird’s DNA, like its Life History. The rhythms are maintained without the stimulation of conditions from the environment. Scientists have learned this by placing birds in a laboratory, maintaining constant light to simulate an unchanging photoperiod, and measuring the pattern of rhythms of their physiological systems. These endogenous rhythms are referred to as free-running.
-
- Free-running circannual rhythms vary among bird species from 9 to 13 months. Even birds in tropical regions, where changes of season are minimal or non-existent, display an endogenous annual cycle.
- Free-running circadian rhythm among birds recurs about every 23-25 hours, depending on the species.
But wild birds do not live in the artificial environment of a laboratory, and their endogenous rhythms are synchronized with and controlled by exogenous systems of the natural world, the daily and annual cycles of the photoperiod. This synchronization is referred to as entrainment. The regular and entirely predictable changes in the photoperiod over the year entrain the endogenous circannual rhythms to the exogenous rhythm of the seasons, so a species’ Life History is repeated within the 365-day timeframe of the Earth’s revolution around the sun. And the endogenous biological clock is entrained to the exogenous changes in the photoperiod over the 24 hours of the Earth’s rotation on its axis. (Scientists sometimes refer to the photoperiod as a Zeitgeber (Ger. Zeit = time + Geber = giver) because of its role in entraining an organism’s circadian rhythms to the Earth’s exogenous rhythms. This compound German word was invented in the 20th century by Jürgen Aschoff (1951), who was a chronobiologist, a scientist who studies biological rhythms.)
HOW DOES PHOTOPERIODISM WORK IN BIRDS?
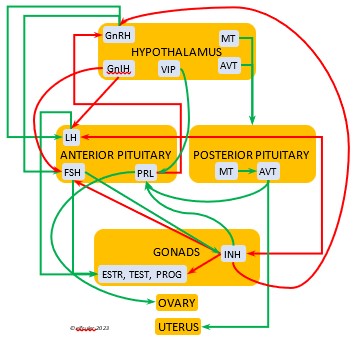
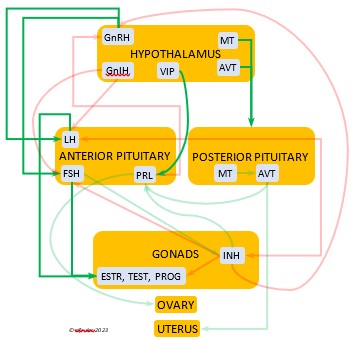
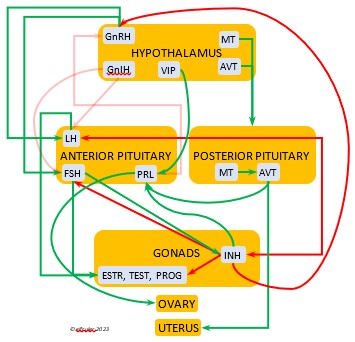
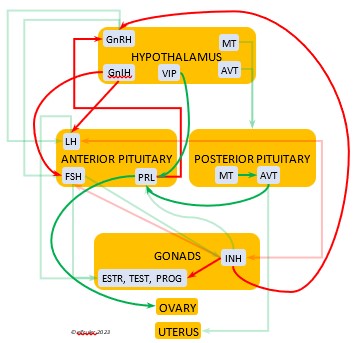
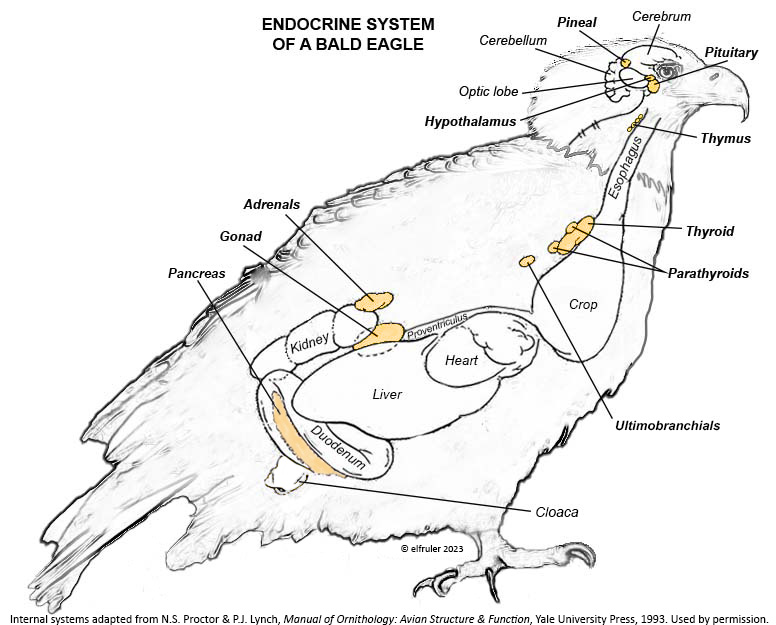
Photoperiodism refers to how an organism responds to the changing photoperiod. The mechanism begins in the brain. The Pineal Gland, the Hypothalamus, and in some species the Retinas of the eyes serve as a bridge between the exogenous world and the endogenous systems of the body. In addition to performing their other functions within the nervous system, these organs are the body’s principal internal oscillators or pacemakers, interacting with each other to maintain a body’s circadian rhythm. The pacemakers also receive information from the external environment by means of photosensitive cells, or photoreceptors. These cells are active during the daylight hours and perceive the presence of sunlight and its intensity and measure the daily photoperiod. They also retain a memory of sunlight changes over the course of the day, and thus the brain can deduce the approximate time of day—early morning, mid-afternoon, etc. (Dawson, King et al. 2001)
As they measure the photoperiod, the photoreceptors stimulate the Pineal Gland to produce the hormone Melatonin (MEL). MEL is inhibited by light, so it is secreted only during the scotoperiod, increasing as daylight diminishes near sunset and decreasing as the sun rises. MEL’s daily rhythmic secretion is the means by which the body’s endogenous daily biological rhythms are synced—or entrained—to the exogenous rhythms of the photoperiod. MEL also induces physiological changes in the body. It signals the brain to decrease its activity during the dark hours, and its circulation through the bloodstream causes lower body temperature and decreased metabolism. (See more about Melatonin, the Pineal Gland, and the Hypothalamus in the article on the Endocrine System and Hormones).
Memory of the daily photoperiod persists throughout the year. The photoreceptors in the brain measure and retain memories of changes in daylight from one day to the next. Thus they can perceive whether the photoperiod is comparatively long or short, whether it is increasing or decreasing, and whether the change is gradual or rapid. The brain also responds to the intensity of sunlight—whether the sun is high overhead or hanging low in the sky (and distorted by atmospheric refraction), as dictated by the time of day and the season. The photoreceptors’ perception of the changes in the photoperiod from one day to the next enables a bird to perceive whether it is spring or fall, summer or winter, in other words to know where they are in their annual Life History and to anticipate what is likely to happen in their environment in coming weeks.
The changing photoperiod is what researchers call a proximate factor, a current or impending situation that triggers behaviorial or physiological responses. The photoperiod is the only proximate factor that is unfailingly consistent from year to year and thus can serve as a signal to the body to prepare for transition to a new Life-History stage. For most birds in the Temperate zone, including all Bald Eagles, the photoperiod is the initial predictive cue that is reliable weeks in advance as a signal to the body to begin the transition from one Life-History stage to the next. (Less predictable proximate factors, such as temperature, food supplies, weather conditions, bonding with a mate, territorial disputes, and human disturbance are discussed below).
Photoperiod affects a bird’s activities and biology throughout its annual cycle, but particularly at two pivotal times for a breeding bird: 1) at the end of the Reproduction stage when the Testes, Ovary, and Oviduct shrink (gonadal regression), and 2) at the start of the next Reproduction stage when the Gonads redevelop (gonadal recrudescence). Gonadal maturity obviously is necessary for the production of sperm and eggs, while gonadal regression precludes the possibility of reproduction during an unfavorable season of the year, and it frees up metabolic resources for Molt and Movement.
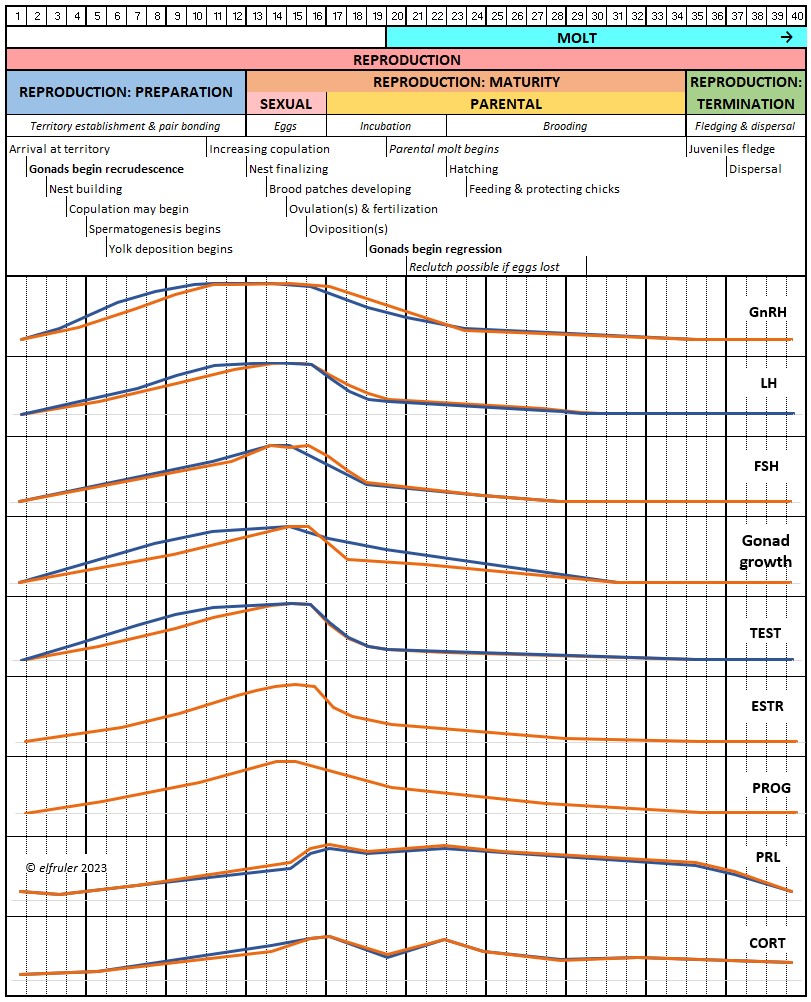
The body’s responses to the changing photoperiod follow a progression of three states for breeding birds (see more detail article on Reproduction and Hormones), which largely coincide with the Life History stages:
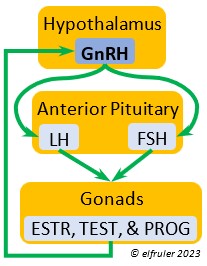
Photosensitivity. In this state the bird becomes sensitive to changes in daylight after a long period of insensitivity and gonadal regression, and the body slowly transitions into the season of Preparation for Reproduction. A protein (opsin) in the brain’s photoreceptor cells undergoes a chemical reaction that signals the Hypothalamus to produce a hormone (GnRH, or Gonadotropin-Releasing Hormone) which initiates a series of hormonal secretions downstream, causing the Gonads begin to regrow. The gonadal recrudescence process is slow, but the hormonal changes gradually affect the birds’ behavior. The Testes begin to produce sperm and yolk material is slowly produced in the liver for deposition in the Ovary. The pair becomes active in their breeding territory, they start to build or refurbish nests, and they engage in bonding behaviors, including copulation.
Photostimulation. As the photoperiod continues to change, the brain’s photoreceptors stimulate significantly increased secretions from the Hypothalamus and the Pituitary Gland which activate sex hormones (Testosterone, Estrogens, and Progesterone) in the Gonads. The Gonads grow rapidly and the bird enters the Maturity phase. Nests are finalized, copulation becomes more frequent, ovulation and fertilization occur, and eggs are laid (oviposition). After oviposition, different hormones induce a transition to a less sexual and more parental phase as adults care for growing nestlings.
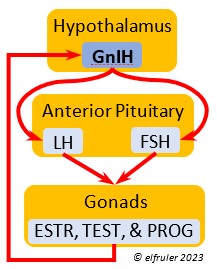
Photorefractoriness. The birds transition into a state of insensitivity to daylight as it settles into a long period of slow daily changes. With little exogenous stimulation and a decline of reproductive hormones, gonadal regression begins and sexual activity between adults recedes as the Termination phase of Reproduction settles in. Chicks grow and ultimately leave their nests. Photorefractoriness continues through the beginning of Molt and the Movement stage of Life History when adults begin migration or other movements outside the immediate nest area. Reproduction will not begin again until Molt is complete and photorefractoriness is dissipated at the beginning of a new Reproduction season.
This schematic diagram illustrates how these photoperiodic states correspond with the annual cycle of a Bald Eagles’ Life-History stages (click on the image for an enlarged view):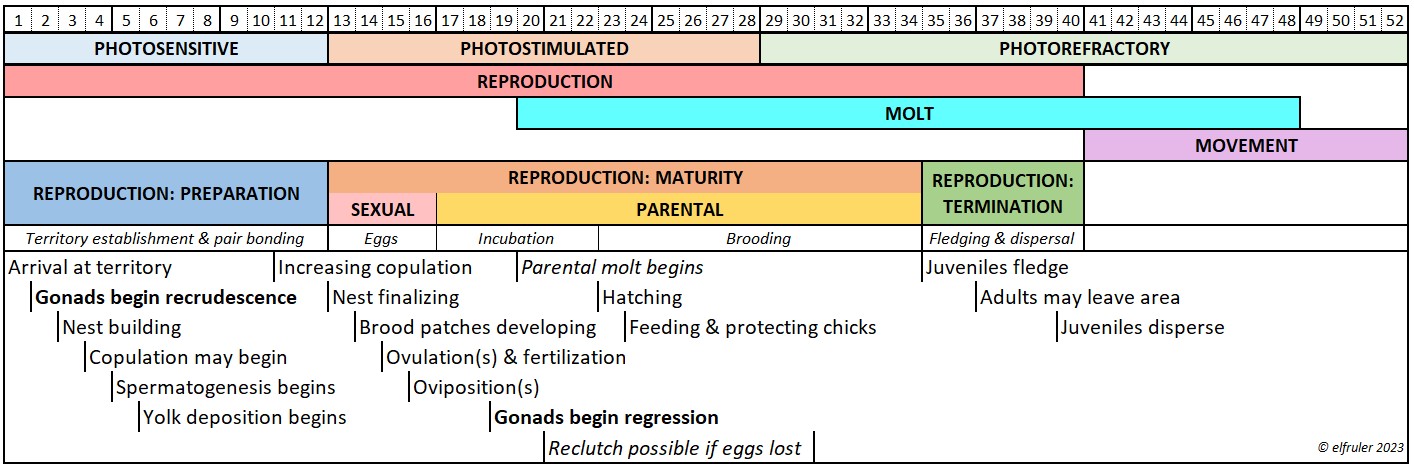
Reproduction occupies well over half of the year for Bald Eagles, and the Preparation phase begins many weeks before they lay eggs. Scientists theorize that the critical predictive cue to the brain is when the daily changing photoperiod reaches a threshold of a certain length, a critical photoperiod. This triggers the dissipation of photorefractoriness and brings the Reproductive system to a state of photosensitivity, when the long slow process of gonadal recrudescence begins. Another critical photoperiod at the end of Reproduction initiates the beginning of photorefractoriness and gonadal regression. What that “threshold of a certain length” is that defines a “critical photoperiod” cannot be pinpointed to a specific day, but it occurs within a brief window of time that maximizes a bird’s likelihood of reproductive success in the weeks that follow. This timeframe differs among species, populations, latitude, and climate, and it can shift for an individual bird from year to year.
ADAPTATION, VARIABILITY, and PLASTICITY
Bald Eagles have a relatively simple Life History (in contrast, for instance, to birds who undergo more than one Molt each year), which enables them to be flexible breeders in the timing of Reproduction, Molt, and Movement, within the boundaries of the genetically established timeframe. (Wingfield 2005) They inhabit most of North America, and the Life History schedules of different populations in this enormous area vary, a type of environmental plasticity. In general the Bald Eagle breeding season occurs progressively later in the year from south to north. But geographical latitude is not the only variable. Geophysical features like coasts, mountains, and deserts can affect climatic conditions (temperature, humidity, rainfall) that shape the timing of breeding. (Dawson 2008) The schedule may depend on habitat, such as whether a pair is nesting in an urban or a rural area. Even within the same region and climate, Bald Eagles’ egg-laying schedules can differ by a month or more, probably dictated by individual genetic make-up, lineage and experiences, and Life History. And although most individuals produce eggs each year within the same timeframe of 1-3 weeks, they may adjust their schedules if necessary due to unforeseen and disruptive events.
Bald Eagles inhabit the large Temperate Climate Zone of North America, from the Sub-Tropics (north of the Tropic of Cancer at ~ 23.44° N) to the Sub-Arctic (south of the Arctic Circle at ~ 66.5° N). This map marks the Tropic of Cancer and the Arctic Circle with thick yellow lines, and I have added red dots showing the locations of Bald Eagle nests recently live-streamed on the internet (click on the image for an enlarged view).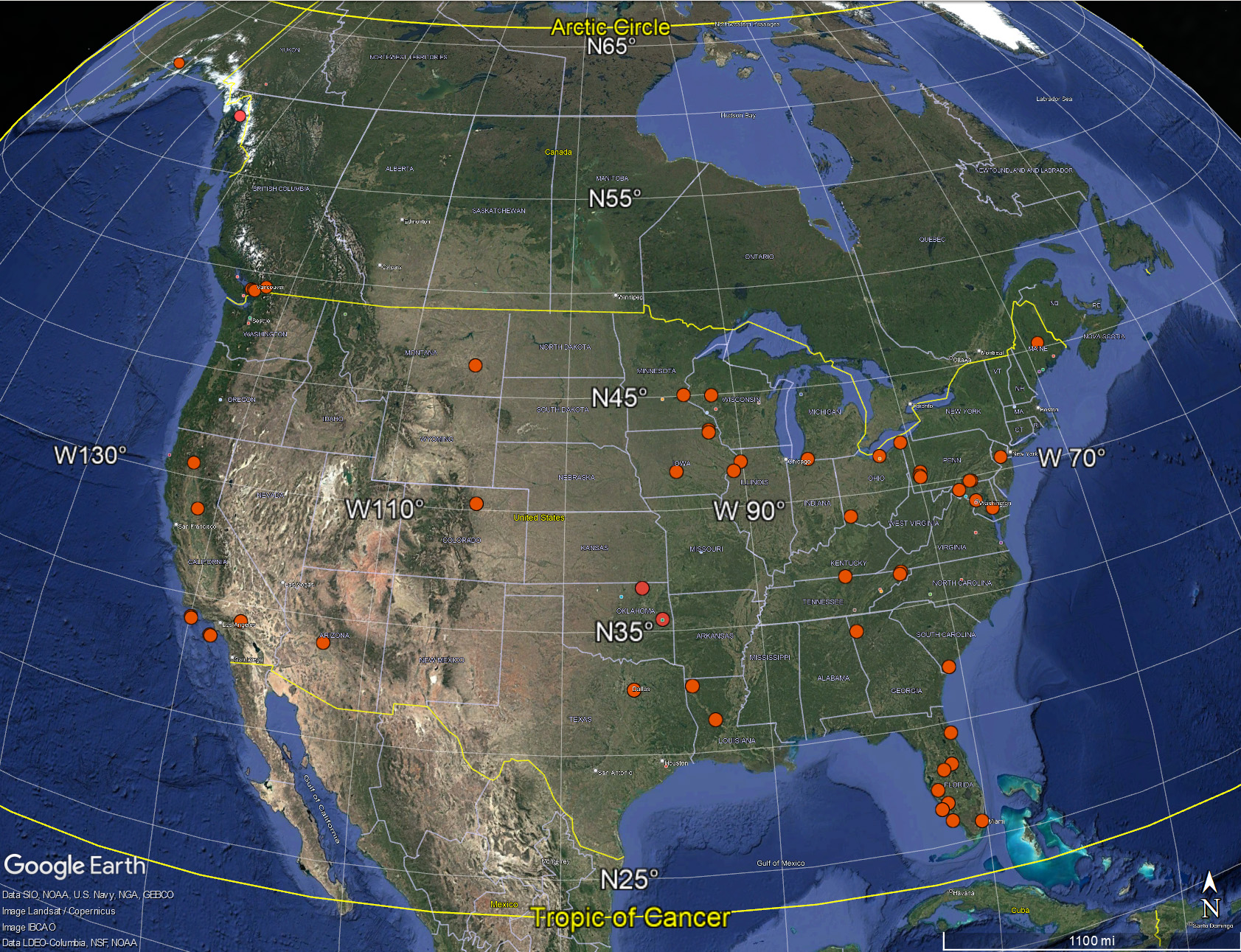
Based on the timing of breeding activities of Bald Eagles, I have separated the Temperate zone into several regions by latitude:
-
- Sub-arctic: just below the Arctic Circle, from about 60° – 50° N, including Alaska and Yukon Territories
- Northern tier: the Northern tier between about 50° – 43° N, including Maine, Massachusetts, Michigan, Wisconsin, Minnesota, Montana, Oregon, Washington, and British Columbia
- Middle tier: between about 43° – 35° N, including Tennessee, North Carolina, Virginia, West Virginia, Maryland, New Jersey, New York, Pennsylvania, Ohio, Indiana, Illinois, Missouri, Iowa, Colorado, Arizona, and California
- Sub-tropics: from about 35° – 23.4° N, including Florida, South Carolina, Georgia, Louisiana, Texas, and Oklahoma
The northernmost nests on cam are those in Alaska at about 60° N, some 448 miles south of the Arctic Circle. The southernmost nest is in south Florida at about 25° N, a few degrees north of the Tropic of Cancer and about 1725 miles north of the Equator.
The different populations of breeding Bald Eagles that these regions delineate remain fairly discrete since eagles tend to return to the general area of their natal nest to establish their own territory. All Bald Eagles have the same Life Histories, but the timing and the progression of the stages are genetically adapted to the climatic conditions of each region, commensurate with timely access to food. Populations at lower latitudes generally begin the Reproduction stage earlier in the year, while those at higher latitudes begin later, a common pattern for wildlife.
The differences among the populations are obvious in the following schematic diagram, which shows the approximate timeframes of the phases and sub-phases of the Reproduction Life-History stage for Bald Eagles from north to south as they have been observed on nest cameras, correlated with changes in photoperiod through the year. (Each Reproduction phase—Preparation, Maturity: Sexual, Maturity: Parental, and Termination—shows the observed time range among all the nests in each region, not the schedule of an individual nest or bird. Click on the image for an enlarged view.)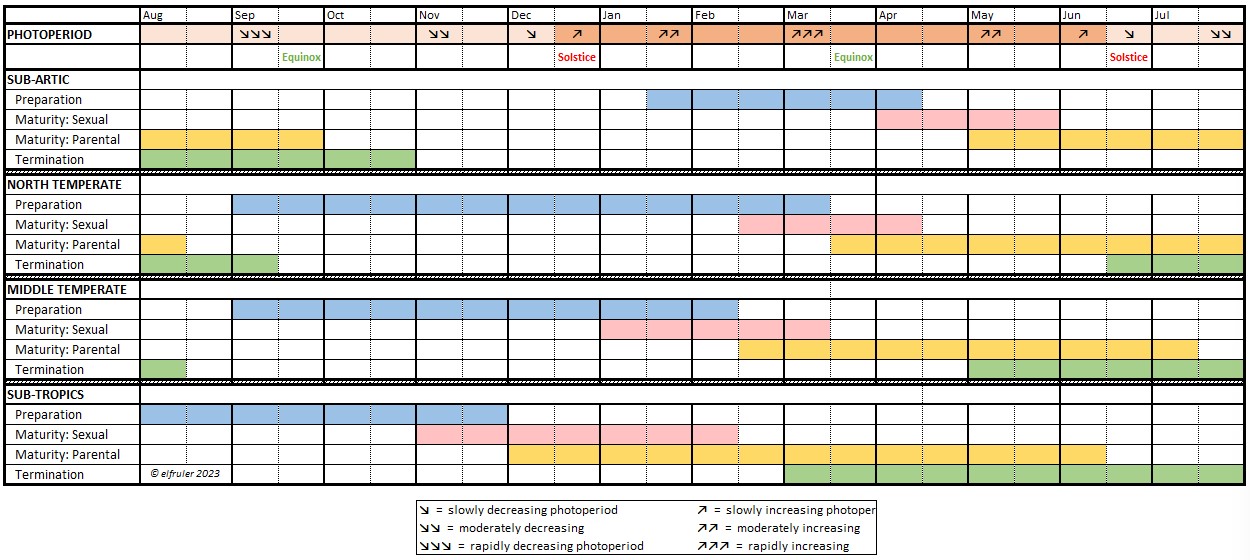
This diagram illustrates a striking contrast in the start of the Reproduction season, the Preparation phase of courtship and nest-building (marked by blue shading in the diagram) when photorefractoriness has dissipated and gonadal recrudescence has commenced. (While we cannot directly see the Gonads of Bald Eagles living in the wild and whether they are growing or shrinking, the eagles’ behaviors reflect the physiological changes.) The difference is whether the eagles start after the Summer Solstice or after the Winter Solstice.
-
- Eagles in the three lower regions (North Temperate, Middle Temperate, and Sub-tropical) begin after the Summer Solstice, from late summer to early fall as daylight is decreasing. Note that eagles in the southernmost Sub-tropics begin only a month or so before the eagles in the North and Middle Temperate regions. This timing is typical of most birds in the Temperate Zone.
- In the extreme northern Sub-arctic region, early breeding behaviors are not evident until several weeks later, when daylight begins lengthening after the Winter Solstice.
A less obvious difference between populations is perhaps more significant: the timing of egg-laying, during the Sexual sub-phase of the Maturity stage (marked by pink shading). The distinction is whether eggs come while the photoperiod is increasing (marked by ↗↗ to ↗↗↗ arrows in the Photoperiod row at the top of the diagram), decreasing (marked by 2 arrows ↘↘ or 3 arrows ↗↗), or barely changing at all (marked by a single ↗ arrow).
-
- Eagles in the northern latitudes (Sub-arctic, North Temperate, and Middle Temperate) lay eggs in late winter and spring when daylight is lengthening more and more rapidly.
- Eagles in the Sub-tropics lay eggs in late fall or early winter as daylight is short or only slowly lengthening.
The early schedule of egg-laying for southern eagles has made some wonder whether photoperiodism plays any role at all in timing their Life History stages. Among scientists there is no real debate about this. While daily changes in the photoperiod in the south are much smaller than further north, studies have shown that birds throughout the Temperate zone and down into the Tropics are highly sensitive to small changes in photoperiod. (Dawson 2007) Like birds further north, birds who breed in the far south have endogenous circadian and circannual rhythms, they experience annual rhythms of gonadal recrudescence and regression as well as Molt, their bodily systems respond to the changing photoperiod, and their endogenous biological clocks are entrained to it. (Gwinner & Scheuerlein 1999) In fact, some birds in the Tropics (well south of the Bald Eagles’ range) can perceive and respond to a difference as small as 17 minutes between the shortest and the longest photoperiod of the year (Hau et al. 1998)—much shorter than the approximately 3.3 hours annual difference for eagles at the southern tip of Florida.
The differences in breeding schedules at different latitudes can be traced to adaptive (genetic) differences in the ways the neuroendocrine systems perceive and respond to the changing photoperiod. (MacDougall-Shackleton & Hahn 2007; Hahn & MacDougall-Shackleton 2008) Some studies suggest that the reproductive systems of populations at lower latitudes may not become completely inactive after the regular breeding season but remain minimally functional and can be reactivated quickly—that their Gonads do not regress completely, but if conditions are favorable they can recrudesce rapidly. (Immelmann 1972; Dawson et al. 2001; Dawson 2008)
Scientists have applied the term long-day breeders to species or populations who lay eggs during the increasingly longer photoperiods in spring and summer, which includes most birds. Experiments have shown that for these birds the initial predictive cue, the critical photoperiod that induces the onset of photosensitivity, occurs when daylight is decreasing after an extended period of long days—i.e. around the Autumnal Equinox. This is the pattern that most Bald Eagles follow. A few species of birds fall into the category of short-day breeders, those who lay eggs as daylight is decreasing or short in the fall or winter. For them, including many southern Bald Eagles, the onset of photosensitivity can occur in the summer while daylight is still long.
Researchers have found that these timing differences between long-day breeders and short-day breeders may be due to how their brains function. (Gwinner & Scheuerlin 1999; Sharp and Blache 2003; Sharp 2005)
-
- In long-day breeders the minimal changes to photoperiod of the long days of summer render the neurons in the Hypothalamus insensitive to daylight. It takes the shift to larger changes in the photoperiod in autumn to “wake up” those neurons so they can begin responding to the changes and send signals to hormones in the Pituitary Gland.
- In short-day breeders the neurons in the Hypothalamus are more active throughout the year and can function even with minimal input from the photoperiod. This means that their reproductive systems may be more readily controlled by their endogenous circannual systems and the neurons can begin to send signals downstream earlier.
(This may help explain events in 2019-2020 at the Southwest Florida eagles’ nest (Fort Myers, FL) where several weeks after losing their 37-day old chick the eagles produced a very late second clutch and raised those eaglets to fledge. A successful second brood after loss of eaglets (as opposed to eggs) from a first brood is a rare occurrence among Bald Eagles, and it may occur only in climates where their reproductive systems are adapted to make it possible.)
NON-PHOTOPERIODIC PROXIMATE FACTORS IN REPRODUCTION
Photoperiod is the only proximate factor that is consistent from day to day and year to year and thus the only one that a bird’s body can rely on as an initial predictive cue. But other proximate factors, which are often unpredictable or irregular, also can affect the timing of Reproduction (Wingfield et al. 2007). Such supplementary information can help fine-tune the timing of Reproduction in short-term situations. Researchers have found that the effect of some proximate exogenous cues on reproductive activities may be more significant in southern birds than in northern birds. (Dawson 2008; Hahn et al. 2009; Silverin et al 2008; Sturkie’s 6th: 504) Non-photoperiodic proximate factors include environmental cues and social cues.
Environmental cues tend to be variable and include climate, weather conditions, and food availability. Unpredictable environmental situations (storms, extreme heat or cold) can be disruptive and can delay or prevent reproductive activities, or even result in loss of a clutch, brood, or nest. Male breeders may be more affected by unfavorable environmental cues than females, since male sperm production may suffer if such conditions are persistent, even before females are ready to ovulate.
A particular region generally has a predictable temperature range, and it is tempting to wonder whether the significant differences in ambient temperatures across the latitudes of the Temperate Zone play a role in the timing of Reproduction. The theory is that the start of breeding is triggered to some degree by the onset of warm weather—earlier in the south and later in the north. But scientists have found little evidence that temperature plays a role as a predictive factor. (Cassone & Kumar 2015) Temperatures from day to day or week to week can make unexpected swings, and there is no way for a wild animal to foresee such swings far enough in advance to begin the long Reproductive process. Once the reproductive process has begun, though, a temporary stretch of unusually low temperatures can slow it down and abnormally high temperatures may speed it up.
Some regions have fairly regular seasons of rainfall, but as with temperature ranges these do not appear to have a predictive impact on the timing of breeding. Unusually low or high rainfall as well as extreme temperature swings can adversely affect the availability of food. Inadequate food in a particular year can stop a breeding season before it starts, and it can induce potential parents to delay or scuttle a breeding attempt, tax the health of breeding adults, or impair the growth and development of nestlings.
The clouds that accompany rainfall may reduce sunlight intensity, the amount of radiation—i.e., energy in the form of light and heat—that reaches the earth by scattering its radiation. Sunlight, of course, is coordinated with the photoperiod throughout the year, but its intensity is dependent on the time of year and time of day. At any given latitude, sunlight intensity is greatest and longest at the Summer Solstice, when it is at its maximum elevation of the year. It is greatest in the middle of the day when the sun is at its highest elevation. It is lowest in early morning and evening both because its lower angle in the sky increases the distance the radiation must travel to reach the earth’s surface, and because at those times its effect is distorted by atmospheric refraction. So if clouds are present at mid-day but build up late in the afternoon, they have less effect on the amount of sunlight intensity reaching the Earth.
Prolonged cloud cover over several days can lower the sun’s intensity, reducing the signals that a bird receives from the photoperiod. Such extended periods can happen almost anywhere, and it may affect birds in the short term, as for instance inducing females to delay ovulation or oviposition. (Dawson 2008) Some areas may experience a regular annual rainy season, suggesting the possibility that it affects the timing of Reproduction. For example, parts of Florida commonly experience a rainy season from June through September. (Daily details of weather features for a given location can be explored here.) Could this be a reliable trigger for gonadal recrudescence and the beginning of Reproduction activities? It is a question impossible to answer with what we know. But while it may be one factor in the timing of Reproduction, a general annual pattern of rain and cloud cover in a particular region can be variable from year to year, day to day, and hour to hour, and the effect on light intensity depends on the time of day that clouds are present. Such variabilities might make cloud cover unlikely to be as reliable a predictive factor every year as the changing photoperiod, although it could have played a role in long-term adaptation to a local climate. (Gwinner 2003; Sturkies 5th, 811; Ronneberg 2010)
Another pertinent point is that while Florida’s summer rains may have played some role in the adaptation of the Life History of the population of Bald Eagles there, a similar situation at comparable latitudes, such as Louisiana, South Carolina, Texas, and Georgia, is not evident. Eagles there have the same egg-laying schedule as Florida but not the same cloud cover pattern, so in those regions photoperiod is the only predictive cue to govern the Reproduction schedule.
Prolonged or persistent exogenous cues, such as climate change, can impact a species or a population over the long term by inducing permanent adaptations that enhance the likelihood of success, and thus become ultimate factors.
Social cues include presence (or absence) of a suitable mate, appropriate interactions between male and female, competition for a mate or a territory, nest site availability, and human disturbance. Pair-bonding and nest-building stimulate hormonal secretions and bring a male and a female into hormonal and behavioral sync, which is crucial to successful breeding (see discussion in the article on Reproduction and Hormones).
Disruptions by intruders or humans are unpredictable and can delay or prevent reproductive activity. Female breeders generally are more affected by social cues such as effective (or inept) courtship behavior by a male. Female Bald Eagles are especially dependent on social cues in the final stages of the Maturity phase of egg production, to induce yolk deposition in the Ovary as well as ovulation. (Ball & Ketterson 2008) But both females and males are affected by other social factors such as intruders, loss of a nest, or human activities.
You must be logged in to post a comment.